Emerging evidence suggests that microbial metabolites not only regulate endogenous GLP-1 secretion but may also influence therapeutic response to GLP-1 receptor agonists, raising the prospect of microbiome-guided treatment strategies in obesity and type 2 diabetes.
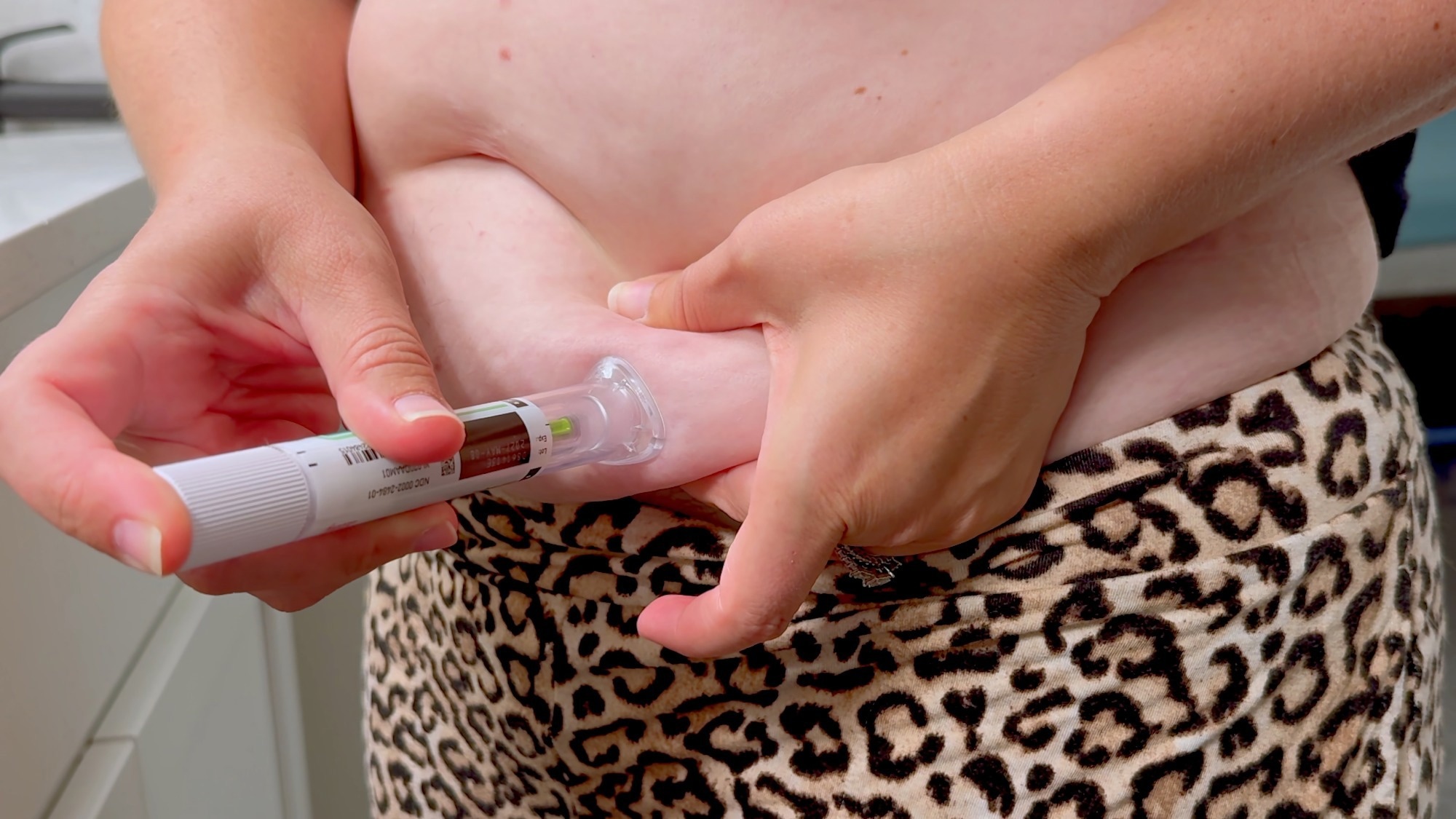
Study: GLP-1 agonists and the gut microbiome: A bidirectional relationship. Image Credit: Matt Fowler KC / Shutterstock
In a recent review published in the British Journal of Clinical Pharmacology, a group of authors examined the bidirectional relationship between glucagon-like peptide-1 receptor agonists (GLP-1RAs) and the gut microbiome and evaluated implications for metabolic health and personalized medicine.
Background
More than 500 million people worldwide live with type 2 diabetes mellitus, and obesity rates continue to rise. GLP-1RAs have improved treatment by improving glucose regulation, supporting weight reduction, and decreasing cardiovascular risk. However, an important unanswered question is why some individuals respond differently to these therapies than others.
New studies suggest the gut microbiome, the trillions of bacteria that live in the gastrointestinal tract, can affect metabolism, immune function, and hormone signaling. Understanding how gut bacteria influence glucagon-like peptide-1 (GLP-1) activity may help improve treatment strategies. However, in the long term, human mechanistic research is still needed.
Microbial regulation of GLP-1 secretion
The gut microbiome helps regulate GLP-1, a hormone secreted by enteroendocrine L-cells in the intestine. GLP-1 stimulates insulin secretion, lowers glucagon, slows gastric emptying, and induces satiety. These actions influence blood glucose and body weight, both of which are central to type 2 diabetes mellitus and obesity.
When gut bacteria break down dietary fiber, they produce short-chain fatty acids (SCFAs), including acetate, propionate, and butyrate. SCFAs activate G-protein-coupled receptors (GPRs) 41 and 43, also known as free fatty acid receptor 3 and free fatty acid receptor 2, on L-cells and engage intracellular signaling pathways. This signaling can stimulate GLP-1 release through mechanisms that include modulation of intracellular calcium and cyclic adenosine monophosphate (cAMP) signaling, as well as indirect gut, brain metabolic pathways described in experimental models. Butyrate can also inhibit histone deacetylase, increasing transcription of the proglucagon gene and supporting ongoing hormone production.
Bile acids also regulate GLP-1. Primary bile acids such as cholic acid and chenodeoxycholic acid are converted by intestinal bacteria into secondary bile acids, such as deoxycholic acid and lithocholic acid. These activate Takeda G-protein receptor 5 (TGR5), increasing cAMP and Epac-mediated vesicle exocytosis and promoting GLP-1 secretion. In contrast, activation of the farnesoid X receptor (FXR) suppresses GLP-1 synthesis, illustrating a dynamic balance shaped by microbial metabolism.
In dysbiosis, increased lipopolysaccharide activates Toll-like receptor 4 (TLR4), triggering nuclear factor kappa B (NF-κB) signaling and the production of inflammatory cytokines, including tumor necrosis factor alpha (TNF-α) and interleukin-6 (IL-6). This inflammation reduces insulin receptor substrate-1 activity and may impair GLP-1 signaling, potentially contributing to reduced drug effectiveness in insulin resistance.
How GLP-1RAs reshape the microbiome?
GLP-1RAs, including liraglutide, semaglutide, and dulaglutide, influence energy intake, bile acid circulation, and intestinal motility in addition to glucose regulation, which can affect microbial composition.
Clinical and preclinical studies report that treatment may enrich beneficial bacteria such as Akkermansia muciniphila and, in some studies, increase Bacteroidetes while reducing Firmicutes or inflammation-associated taxa. Some trials report increased microbial alpha diversity and beta diversity, whereas others find minimal shifts, suggesting variability between individuals and differences in study design, duration, diet, and concomitant therapies.
In patients with type 2 diabetes mellitus, liraglutide has been associated with lower glycated hemoglobin (HbA1c) and modest changes in body weight and body mass index (BMI), alongside increased abundance of Akkermansia muciniphila. In high-fat diet animal models, semaglutide has been associated with a reduced Firmicutes-to-Bacteroidetes ratio, lower inflammatory cytokines, and improved glucose tolerance.
Microbial changes may not reflect only direct drug effects. Reduced caloric intake and weight loss can independently alter microbial communities, and medications such as metformin also reshape the microbiome, complicating interpretation. These confounders highlight the need for controlled longitudinal studies with standardized dietary monitoring and, where feasible, metabolomic profiling.
Metabolic implications beyond glucose control
Beyond glucose control, GLP-1RAs may affect liver triglyceride levels, fat metabolism, and the conversion of white adipose tissue into brown adipose tissue, as shown in animal studies. Semaglutide has been associated with increased expression of genes involved in adenosine monophosphate-activated protein kinase (AMPK) signaling and carnitine palmitoyltransferase 1 (CPT1).
Fecal microbiota transplantation experiments suggest that transferring microbiota from treated animals to antibiotic-treated recipients can reproduce improvements in weight and insulin sensitivity, indicating microbial shifts may be functionally relevant rather than incidental.
For individuals using these therapies for weight loss, microbial contributions could help explain sustained benefits even when dietary patterns fluctuate. However, human causal data remain limited, and definitive fecal microbiota transplantation studies in patients are lacking.
Precision medicine and predictive potential
Variability in treatment response remains a major clinical challenge. In a pilot study of 52 individuals with type 2 diabetes mellitus, responders to GLP-1RAs had distinct baseline microbial signatures compared with nonresponders. Species such as Bacteroides dorei and Roseburia inulinivorans were associated with greater reductions in HbA1c.
Machine learning models using microbial features achieved promising, though preliminary, predictive accuracy in small cohorts, suggesting that microbiome profiling could support more personalized therapy selection.
Multiomics approaches and gut-on-a-chip microfluidic systems may clarify whether modifying microbial composition can enhance drug efficacy or reduce variability, potentially supporting tailored therapy rather than trial-and-error prescribing.
Conclusions
Current evidence suggests that GLP-1RAs and the gut microbiome influence one another through metabolic and inflammatory pathways. Microbial metabolites regulate GLP-1 secretion, while GLP-1RAs may reshape microbial composition and potentially amplify metabolic benefits. Although preclinical findings suggest functional relevance, causal evidence in humans remains limited. Microbial profiles may partly explain differences in treatment response, supporting interest in personalized medicine. Future research should include controlled dietary parameters, longitudinal follow-up, and microbiome-targeted interventions to better define host-microbe-drug interactions and improve long-term outcomes in the management of obesity and type 2 diabetes.
 Oral microbiome imbalances may contribute to symptomatic hand arthritis
Oral microbiome imbalances may contribute to symptomatic hand arthritis